Autores: Luiz Eduardo Braga¹, Eduardo Argenta Steinhaus¹, Camila Tenório¹, Djordan Willian Feller¹, Cledison Conte² e Marcelo Gripa Madalosso³
Considerada uma das doenças da soja de maior severidade no mundo, a ferrugem asiática, ocasionada pelo agente etiológico Phakopsora pachyrhizi, ocorre na maioria das regiões produtoras de soja do Brasil e do mundo, principalmente nas Américas, Ásia e África (YORINORI et al., 2004). Existem diversos fatores que fazem com que este patógeno possua tamanha importância no sistema de produção de soja, seja pela capacidade de causar perdas de produtividade, pela alta produção de esporos, que resulta em maior ocorrência da doença no campo e junto do controle químico, o aparecimento de resistência aos fungicidas, pelo uso de outras espécies vegetais como hospedeiros alternativos e também pela facilidade de dispersão (SINCLAIR & HARTMAN, 1999; REIS et al., 2018; RYTTER et al., 1984; FANARO et al., 2004).
Por ser um patógeno biotrófico, seus esporos possuem grande capacidade de disseminação por correntes aéreas, atingindo uma grande extensão territorial. Desta forma, o entendimento da dinâmica de dispersão do esporo da ferrugem asiática, passa a ser uma alternativa importante para o auxílio no manejo da doença, pois pode possibilitar uma previsibilidade na chegada do patógeno na área, sua evolução e consequente epidemia. Solidificando estas informações, as tomadas de decisão dos manejos agronômicos atingirão melhor sucesso e reduzirão os danos ao meio ambiente.
O estudo da epidemiologia do patógeno é um fator básico para compreender como a doença ocorre no campo. Dentro da epidemiologia de P. pachyrhizi, a fase aerobiológica evidencia como os esporos se dispersam no ambiente e como os fatores climáticos influenciam o processo. O processo de disseminação dos esporos de P. pachyrhizi engloba as fases de soltura dos esporos que estavam nas urédias, diluição dos esporos na atmosfera, sobrevivência dos esporos na atmosfera e deposição dos esporos em superfícies vegetais ou não (DEL PONTE & ESKER, 2008).
A soltura dos esporos produzidos pelas urédias no ambiente pode ocorrer pela ação do vento movimentando o dossel ou das gotas de chuva que caem sobre a superfície das plantas, já que o patógeno não possui um mecanismo específico de liberação, onde esses esporos podem ser dispersos sozinhos ou em massas de esporos (WALLACE & HOBBS, 1977; MELCHING et al., 1979; DEL PONTE et al., 2006).
A diluição dos esporos na atmosfera ocorre apenas com uma baixa porcentagem dos esporos produzidos, onde são carregados principalmente por difusão turbulenta e cisalhamento do vento, transformando-se em aerossóis, estando dispersos em matéria seca ou líquida no ar, sendo que a distância percorrida pelos esporos é variável de poucos quilômetros até distâncias continentais, o que depende de uma série de fatores climáticos vigentes no momento da disseminação (LI et al., 2009; ISARD et al., 2005). As fortes chuvas servem como veículo para o transporte dos esporos que estão na atmosfera até a superfície, servindo não só como forma de disseminação do patógeno, mas também, como fonte de umidade para o início da infecção (PAN et al., 2006; LI et al., 2009; NAGARAJAN & SINGH, 1990).
A sobrevivência dos esporos na atmosfera é dependente de fatores do patógeno e do ambiente. De maneira geral, os esporos de P. pachyrhizi possuem poucas estratégias de se manter viáveis no ambiente, sendo que a formação de massas de esporos talvez seja o mais importante (Figura 1). O aglutinamento de esporos confere proteção física a aqueles esporos que estão localizados no interior do aglomerado, sendo que os esporos localizados na superfície externa da massa possivelmente serão mortos pelos efeitos da radiação solar, da baixa umidade e da alta temperatura (MELCHING et al., 1979; KIM et al., 2005) (Figura 2).


A ocorrência de nuvens de tempestades, como as que se formam em furacões, possuem a capacidade de proteger os esporos de P. pachyrhizi contra a radiação UV (PAN et al., 2006).
Os fatores ambientais limitantes a viabilidade dos esporos são principalmente temperatura, radiação solar e umidade relativa, onde radiação solar igual ou superior a 27,3 MJ/m², temperatura do ar acima de 30ºC e umidade relativa do ar abaixo de 40% reduzem drasticamente ou inibem a germinação dos esporos (HEGDE et al., 2002; ISARD et al., 2006).

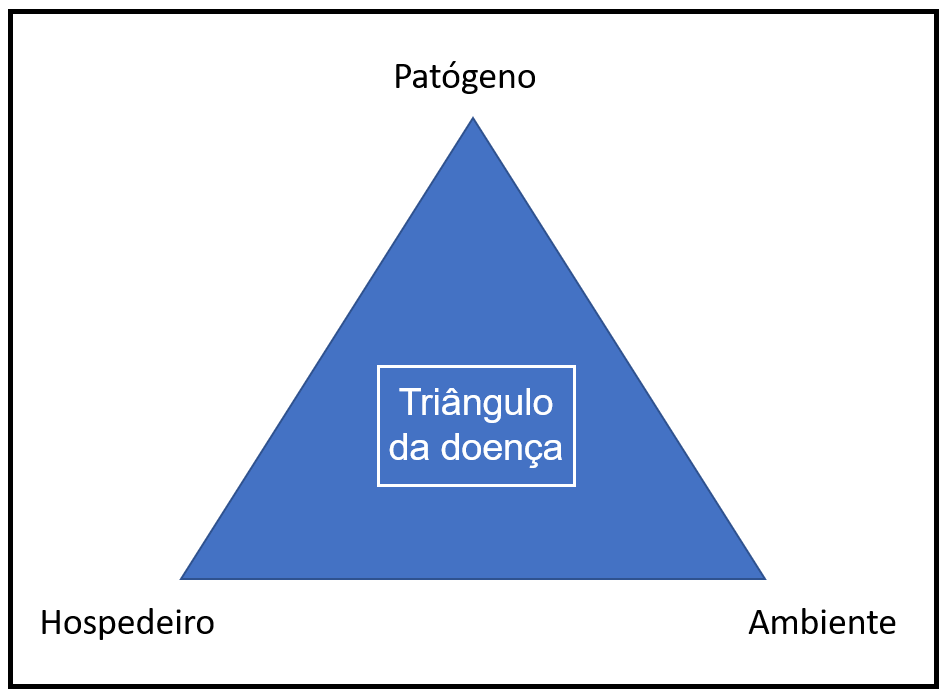
Por último, a deposição dos esporos após a disseminação ocorre em qualquer superfície, podendo ser no solo ou em superfícies vegetais. Após a deposição do esporo, a germinação deste poderá ocorre mesmo em outras superfícies que não sejam tecido vegetal, se este esporo for viável e as condições de umidade e temperatura forem favoráveis (AYLOR, 1986; DEL PONTE & ESKER, 2008). Entretanto, por se trator de um patógeno biotrófico obrigatório, a infecção no tecido vegetal só ocorrerá nas plantas hospedeiras do patógeno se todas as condições do triângulo de ocorrência da doença (figura 3) (hospedeiro suscetível, patógeno viável e ambiente favorável) estiverem ocorrendo (ALVES et al., 2006; MELCHING & BROMFIELD, 1975).
Caracterizando a movimentação de esporos de P. pachyrhizi nos países produtores de soja na América do Sul, alguns fenômenos atmosféricos podem estar associados. O fenômeno dos jatos de baixos níveis da América do Sul (South American Low-Level Jet ou SALLJ) é um corredor de umidade formado pelos ventos alísios que entram pelo nordeste brasileiro e se deslocam pela Amazônia até os Andes, onde as montanhas servem como barreiras, criando corredores de umidade que se deslocam em direção ao sul, passando pela Bolívia, Paraguai, centro-oeste, sudeste e sul do Brasil, chegando no Uruguai e Argentina (figura 4), principalmente durante o verão (MARENGO et al., 2004).
Esse corredor de umidade além de ser responsável pela distribuição de chuvas no continente, também é responsável pela movimentação de partículas no sentido norte-sul, como poeiras e fumaça.
Independentemente do processo de formação, a chuva é a principal responsável pela limpeza da atmosfera, carregando de volta para a superfície todas as partículas que ficam em suspensão, incluindo os esporos. A água da chuva também é interceptada pelas folhas das plantas e fornece, juntamente com a água formada pelo orvalho, o molhamento necessário para a germinação e infecção pelos esporos.
Por outro lado, a chuva também é responsável pelo impacto da gota na folha e a liberação dos esporos da pústula. Após isso, o vento fica responsável pelo deslocamento a longas distâncias. Em períodos secos, após a formação da pústula, a liberação e mobilidade do patógeno são dificultadas. Por isso que, após as precipitações, os componentes climáticos e fitopatológicos se somam para potencializar a epidemia.
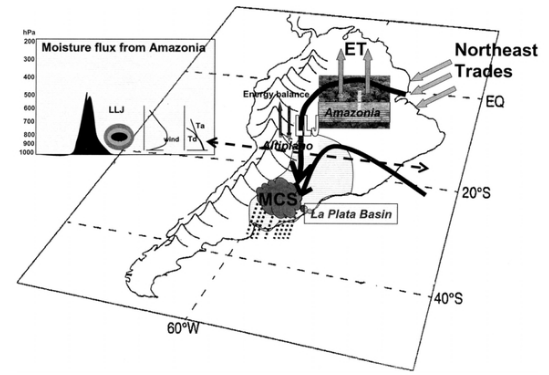
Sendo assim, pode-se relacionar o trajeto percorrido pelos SALLJ com a distribuição espacial dos esporos de P. pachyrhizi nos estados brasileiros e nos países vizinhos, visto que de maneira geral, os primeiros casos de ferrugem asiática e presença de esporos são observados no centro-oeste brasileiro e em países como a Bolívia, e gradativamente são observados no Paraguai, estados do Sul brasileiro, e por último na Argentina e Uruguai.
Além disso, é importante ressaltar que o coletor de esporos é uma ferramenta para estudarmos dispersão do patógeno e formação de um banco de dados que, futuramente podem ajudar nas aplicações.
Iniciar, parar ou aumentar os intervalos de aplicação com base no coletor de esporos é obra do acaso e não tem segurança técnica.
Portanto, o conhecimento da aerobiologia de esporos de P. pachyrhizi, permite relacionar o nível de coleta dos esporos com as condições meteorológicas de determinada região. Assim, futuramente esse banco de dados poderá servir de ferramenta de manejo para antecipar o manejo desta que é uma das doenças que mais acarreta prejuízos ao produtor rural.
Referências
ALVES, S. A. M.; FURTADO, G. Q.; BERGAMIN FILHO, A. Influência das condições climáticas sobre a ferrugem da soja. In: ZAMBOLIM, L. (Org.) Ferrugem asiática da soja. Visconde do Rio Branco: Suprema Gráfica e Editora, 2006. p.37-59.
AYLOR, D.E. A framework for examining inter-regional aerial transport of fungal spores. Agricultural and Forest Meteorology, v.38, p.263–288, 1986.
DEL PONTE, E. M.; GODOY, C. V.; LI, X.; YANG, X. B. Predicting severity of Asian soybean rust with empirical rainfall models. Phytopathology, v.96, p.797-803, 2006.
DEL PONTE, E. M.; ESKER, P. D. Meteorological factors and Asian soybean rust epidemics: a systems approach and implications for risk assessment. Sci. agric. (Piracicaba, Braz.) [online]. Vol.65, pp.88-97, 2008.
FANARO, G. B.; GUEDES, R. L.; CREDE, R. G.; SABUNDIJAN, I. T.; CLAUDIO, T. B.; BALDASSO, J. G.; GREINER, R.; VILLAVICENCIO, A. L. C. H. “Detection of Phakopsora pachyrhyzi by Polymerase chain reaction (PCR) after E.Beam processim to preserve Soya beans”, In: EFFoST- Food Innovations for an Expanding Europe, Poster abstracts. Warsaw, Polônia, p.2.29, 2004.
HEGDE, A. K. H.; ANAHOSUR, K. S.; KACHAPUR, M. R. Germination of Uredospores of Phakopsora pachyrhizi causing soybean rust. Plant Pathology, v.20, p-24-25, 2002.
ISARD, S. A.; GAGE, S. H.; COMTOIS, P.; RUSSO, J. M. Principles of the atmospheric pathway for invasive species applied to soybean rust. Bioscience, v.55, p.851-861, 2005.
ISARD, S. A.; DUFAULT, N. S.; MILES, M. R.; HARTMAN, G. L.; RUSSO, J. M.; DE WOLF, E. D.; MOREL, W. The effect of solar irradiance on the mortality, of Phakopsora pachyrhizi urediniospores. Plant disease , v.90, p. 941-945, 2006.
KIM, K. S., YANG, X. B., PAN, Z.; PEREZ-HERNANDES, O. Simulation of spore trajectories to identify potential pathways of soybean rust in the United States. Phytopathology 95:S54. 2005
LI, X.; YANG, X.; MO, J. Estimation of soybean rust uredospore terminal velocity, dry deposition, and the wet deposition associated with rainfall. Eur J Plant Pathol. P.123-377, 2009.
MARENGO, J.; SOARES, W.; SAULO, C.; NICOLINI, Nicolini, M. Climatology of the Low-Level Jet East of the Andes as Derived from the NCEP–NCAR Reanalyses: Characteristics and Temporal Variability. Journal of Climate, 17(12), 2261-2280, 2004.
MELCHING, J. S.; BROMFIELD, K. R. Factors influencing spore germination and infection by Phakpsora pachyrhizi and intensification and spread of soybean rust under controlled conditions. Proceedings of the American Phytopathological Society, v.2, p.125, 1975.
MELCHING, J. S.; BROMFIELD, K. R.; KINGSOLVER, C. H. Infection, colonization, and uredospore production on Wayne soybean by four cultures of Phakopsora pachyrhizi, the cause of soybean rust. Phytopathology, v.69, p.1262-1265, 1979.
NAGARAJAN, S.; SINGH, D. V. Long-distance dispersion of rust pathogens. Annu Rev Phytopathol. P. 28:139-53, 1990.
PAN, Z.; YANG, X. B.; PIVONIA, S.; XUE, L.; PASKEN, R.; ROADS, J. Long-term prediction of soybean rust entry into the continental United States. Plant Dis. 90:840-846. 2006.
REIS, E. M.; REIS, A. C.; ZANATTA, M. Reflexo econômico e desenvolvimento da resistência de Phakopsora pachyrhizi a fungicidas em função do número de aplicações. Summa phytopathol. Vol.44, n.3, pp.289-292, 2018.
RYTTER, J. L., DOWLER, W. M.; BROMFIELD, K. R. Additional alternative hosts of Phakopsora pachyrhizi, causal agent of soybean rust. Plant Disease 68:818-819. 1984.
SINCLAIR, J. B.; HARTMAN, G. L. Soybean rust. In: HARTMAN, G. L.; SINCLAIR, J. B.; RUPE, J. C. (Eds.). Compendium of soybean diseases. 4. ed. Saint Paul MN. APS Press. pp. 25-26. 1999.
WALLACE, J. M.; HOBBS, P. V. Atmospheric Science: An Introductory Survey. New York: Academic, 1977.
YORINORI, J. T.; NUNES JÚNIOR, J.; LAZZAROTTO, J. J. Ferrugem “asiática” da soja no Brasil: evolução, importância econômica e controle. Documentos 247. Embrapa Soja. Londrina. 36 p. 2004.
Informações sobre os Autores:
- ¹ – Estudante do curso de agronomia URI Campus Santo Ângelo e membro do Grupo de Proteção de Plantas URI Santo Ângelo.
- ² – Engenheiro Agrônomo – Copronar/Py.
- ³ – Professor Dr. da URI Campus Santo Ângelo e Santiago e coordenador do grupo de proteção de plantas URI Santo Ângelo e Santiago.




{kind=link}