Autores: Eduardo Argenta Steinhaus¹, Luiz Eduardo Braga¹, Gabriel Naresi Roos², Fabrício C. Dalenogare2, Lucas Fernandes2, Thuanny Couceiro2 e Marcelo Gripa Madalosso³.
Os primeiros relatos sobre a ocorrência de ferrugem-asiática (Phakopsora pachyrhizi) no continente americano ocorreu no Paraguai, em março de 2001 (MOREL PAIVA, 2001), alguns meses depois foi contatada a sua presença no Estado do Paraná, no mês de maio, infectando soja (Glycine max (L.)) de segundo cultivo (YORINORI e LAZZAROTTO, 2004).
As condições ótimas para a infeção e desenvolvimento da P. pachyrhizi é a ocorrência de molhamento foliar de no mínimo 6 h, e temperaturas entre 15oC a 25oC (CABI, 2021). De acordo com as condições exigidas no triangulo da doença, o terço inferior é o preferencial para a ocorrência e visualização da ferrugem da soja (Figura 1).
No estágio inicial da infecção, não é possível visualizar a fase latente, onde o patógeno esta infectando e colonizando os tecidos do hospedeiro, antes da esporulação (Figura 2). Em seguida, se observa pequenos pontos escuros, de esverdeados ao cinza esverdeado, este é o momento que o patógeno manifesta sua primeira fase de infecção, que pode ser observado apenas com o auxílio de equipamentos como microscópio (Figura 3).
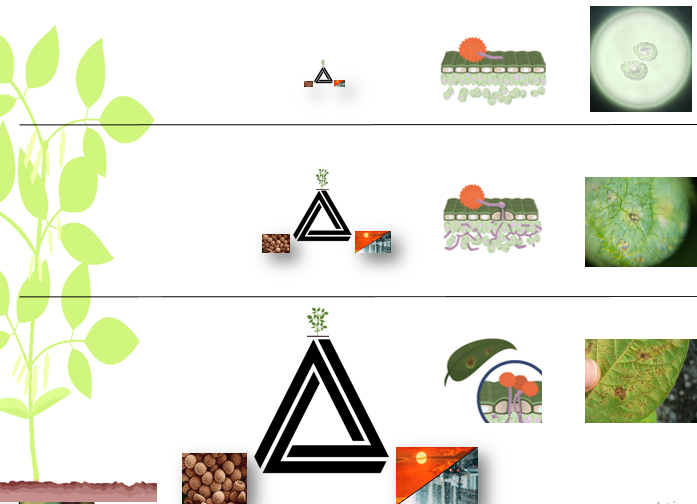

Os sintomas e sinais são encontrados mais comumente nas folhas, entretanto este fungo pode infectar pecíolos e hastes, nesta ordem respectivamente. Os sinais e sintomas podem ser visíveis em todos os estágios da soja, desde que se tenha ambiente adequado para o fungo se desenvolver. No entanto, a presença deles predomina nos estádios reprodutivos da planta.
Existem três tipos de reações percebidas na soja, causadas por P. pachyrhizi (Bromfield & Hartwig, 1980), a primeira é chamada de resistência completa ou (IM) imune, onde o fungo não apresenta o desenvolvimento das estruturas reprodutivas visíveis em seu hospedeiro, tais como, urédias e uredinosporos (Figura 4A).

A segunda lesão é chamada de resistência parcial ou incompleta, que causa o aparecimento e desenvolvimento das lesões do tipo RB (Reddish Brown), causam manchas avermelhadas-escuras, que apresentam baixa taxa de reprodução deste patógeno no hospedeiro, entretanto, o período de latência das urédias é maior que a do tipo TAN (Figura 4B).
A lesão RB ocorre comumente em cultivares com a tecnologia Inox®, ao ponto que quando o hospedeiro percebe a presença do patógeno, ele faz uma destruição localizada das células do tecido vegetal, para evitar o prosseguimento da infecção pelo fungo, esta reação é chamada de hipersensibilidade (Figura 5).
A terceira lesão é chamada de TAN, apresenta pequenas manchas ou lesões de morrom-amarelada ou morrom-acinzentada, delimitadas pelos feixes vasculares. Este tipo de lesão exibe uma intensa esporulação de urédias (Figura 4C). A planta não apresenta nenhum tipo de reação para se defender do fungo.


Tanto a reação IM quanto a reação RB são iniciadas com a percepção precoce do proteínas de avirulência de patógenos por proteínas R vegetais, de acordo com o clássico teoria da resistência gene a gene. Tal teoria diz que para cada gene de resistência constatado no hospedeiro, existe também um gene complementar do patógeno que condiciona a avirulência (FLOR, 1956).
As urédias ou pústulas da doença são visíveis após sua esporulação encerrando assim seu primeiro ciclo. Na parte abaxial da folha forma-se urédias onde ocorre sua disseminação, a coloração das mesmas é marrom escura. Aumentando o nível da doença ocorre a necrose de os tecidos foliares e consequentemente a desfolha precoce (Figura 6). Em casos severos, é possível observar algumas pústulas na porção adaxial da folha (Figura 7). O resultado final é o aborto de vagens, interrupção do enchimento dos grãos e gradualmente perda do potencial produtivo (Figura 8).



Informações sobre os Autores:
- ¹ Estudante do curso de agronomia URI Campus Santo Ângelo e membro do grupo de proteção de plantas URI Santo Ângelo.
- ² Estudante do curso de agronomia URI Campus Santiago e membro do grupo de proteção de plantas URI Santiago.
- ³ Professor Dr. da URI Campus Santo Ângelo e Santiago e coordenador do grupo de proteção de plantas URI Santo Ângelo e Santiago.
Referências Bibliográficas:
BROMFIELD, K. R., and HARTWIG, E. E. Resistance to soybean rust and mode of inheritance. Crop Sci. 20:254-255. 1980.
CABI, 2021. Link: (https://www.cabi.org/isc/datasheet/40019#tosymptoms). Acessado 01/05/2021.
FLOR, H.H. The complementary genic systems in flax and flax rust. Advances in Genetics 8:29-54. Academic Press, New York. 1956.
MOREL PAIVA, W. Roya de Ia soja. Itapúa: Ministério de Agricultura y Ganaderia, Subsecretaria de Agricultura, Dirección de Investigación Agrícola: Centro Regional de Investigación Agrícola – CRIA, 2001. (Comunicado Técnico – Reporte Oficial, Série Fitopatologia, 1).
ROSA C.R.E., SPEHAR CR, LIU J.Q. (2015) Asian Soybean Rust Resistance: An Overview. J Plant Pathol Microb 6: 307.
YORINORI, J.T.; LAZZAROTTO, J.J.. Situação da ferrugem asiática da soja no Brasil e na América do Sul. Embrapa Soja-Documentos (INFOTECA-E), 2004.
Foto de capa: Madalosso, M.G. 2017.




{kind=link}